Jaga:



Liitude rakutüübid ja nende omadused
The raku ristmikud need on kontaktisildad, mis paiknevad külgnevate rakkude vaheliste tsütoplasma membraanide vahel või raku ja maatriksi vahel. Liigud sõltuvad uuritud koe liigist, rõhutades seoseid epiteel-, lihas- ja närvirakkude vahel.
Rakkudes on nende vahelise adhesiooniga seotud molekulid. Siiski on vaja täiendavaid elemente, mis suurendavad kudede sidumise stabiilsust. See saavutatakse rakuliste ristmikega.
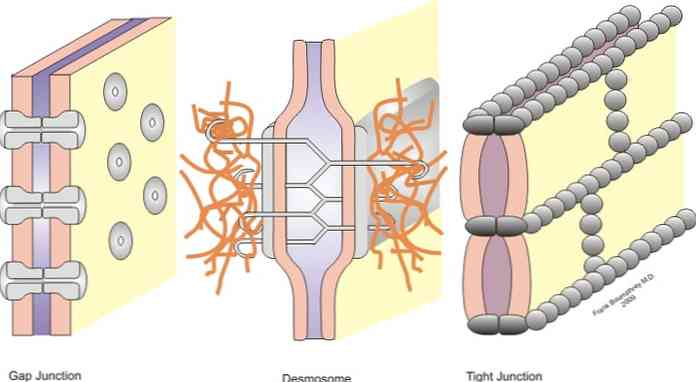
Liiged liigitatakse sümmeetrilistesse liigenditesse (kitsad liigesed, desmosoomid turvavöös ja piluühendused) ja asümmeetrilised liigesed (hemidesmosoomid).
Kitsad ristmikud, rihma desmosoomid, punktidosoomid ja hemidesmosoomid on liigesed, mis võimaldavad ankurdamist; arvestades, et pilu ristmikud käituvad naaberrakkude vahelise ühenduse sildadena, võimaldades soliidide vahetamist tsütoplasmade vahel;.
Lahustuvate ainete, vee ja ioonide liikumine toimub üksikute rakuliste komponentide kaudu ja nende vahel. Seega on olemas transcellulaarne rada, mida juhib mitmed kanalid ja konveierid. Erinevalt paratsellulaarsest rajast, mida reguleerivad rakkude vahelised kontaktid - see tähendab raku ristmikud.
Taimedes leiame rakulisi ristmikke, mis sarnanevad piluühendustega, mida nimetatakse plasmodesmseks. Kuigi need erinevad oma struktuuri poolest, on funktsioon sama.
Meditsiinilisest vaatenurgast tulenevad raku ristmike teatavad puudused omandatud või pärilikest haigustest, mis on põhjustatud epiteeli barjääri kahjustamisest..
Indeks
- 1 Omadused
- 2 tüüpi
- 2.1 - kitsad ühendused
- 2.2 - pilu või pilu ristmikud
- 2.3 - kinnitus- või kinnitusühendused
- 2.4 -Hemidesmosoomid
- 3 Rakkude ristmikud taimedes
- 4 Meditsiiniline perspektiiv
- 5 Viited
Omadused
Elusorganismid koosnevad diskreetsetest ja mitmekesistest struktuuridest, mida nimetatakse rakkudeks. Neid piirab plasmamembraan, mis hoiab neid ekstratsellulaarsest keskkonnast eraldatud.
Kuigi nad on elusolendite komponendid, ei sarnane nad tellistega, kuna nad ei ole üksteisest isoleeritud.
Rakud on omavahel ja ekstratsellulaarse keskkonnaga suhtlevad elemendid. Seetõttu peab rakkudel olema kudede moodustamiseks ja suhtlemiseks, kuid membraan jääb puutumata.
See probleem on lahendatud tänu epiteelil esinevatele rakuühendustele. Need ühendused moodustuvad kahe külgneva raku vahel, liigitatakse vastavalt iga sümmeetrilise ja asümmeetrilise liigese funktsioonile..
Asümmeetrilised ühendused kuuluvad hemidesmosoomidesse ja sümmeetrilistes ühendustes on tihedad ühendused, desmosoom turvavöödel, desmosoomid ja piluühendused. Järgnevalt kirjeldame üksikasjalikult iga ametiühingut.
Tüübid
-Kitsad ristmikud

Kitsad ristmikud, mis on kirjanduses tuntud ka oklusiaalsete ristmikena, on naaberrakkude rakumembraanide sektorid, mis on tihedalt seotud - nimega "kitsas ristmik" näitab.
Keskmistel tingimustel eraldatakse rakud 10 kuni 20 nm kaugusel. Kitsate ristmike korral väheneb see kaugus märkimisväärselt ja mõlema raku membraanid puudutatakse või isegi ühendatakse.
Tüüpiline kitsas ristmik asub naaberrakkude külgseinte vahel nende apikaalsetest pindadest minimaalsel kaugusel.
Epiteelkoes moodustavad kõik rakud sellist tüüpi liite, et jääda ühtseks. Selles interaktsioonis paiknevad rakud, mis moodustavad ringi meenutava mustriga. Need ametiühingud katavad kogu perimeetri.
Valgud, mis on seotud tihedate ristmikega
Ocludina ja Claudina
Tiheda kontakti piirkonnad ümbritsevad kogu raku pinda. Need piirkonnad moodustavad transmembraansete valkude, mis on tuntud kui okstludiin ja claudin, anastomootsed kontaktribad. Termin anastomoos viitab teatud anatoomiliste elementide liitumisele.
Need kaks valku kuuluvad tetraespaniinide rühma. Neile on iseloomulik, et neil on neli transmembraanset domeeni, kaks välist silmust ja kaks suhteliselt lühikest tsütoplasmaatilist saba.
On täheldatud, et okklusiin interakteerub nelja teise valgumolekuliga, mida nimetatakse zonula oklusiiniks ja lühendatult ZO-ks. See viimane rühm hõlmab valke ZO1, ZO2, ZO3 ja afuna.
Claudin on seevastu 16 valgu perekond, mis moodustab kitsastes ristmikutes lineaarsete fibrillide seeria, mis võimaldab sellel liidul võtta "barjääri" rolli paratsellulaarses rada..
Nectinas ja JAM
Kitsad ristmikud ilmuvad ka nektiinide ja liitude adhesioonimolekulide vahel (lühendatud lühendiga inglise keeles JAM). Need kaks molekuli on rakusiseses ruumis homodimeeridena.
Nektiinid on aktiinfilamentidega seotud afadiinvalgu abil. Viimane näib olevat eluliselt tähtis, kuna närilistel afadiini kodeeriva geeni deletsioonides põhjustavad nad embrüo surma.
Kitsate ristmike funktsioonid
Sellised raku-raku ristmikud täidavad kahte olulist funktsiooni. Esimene on epiteeli rakkude polaarsuse määramine, apikaalse domeeni eraldamine basolateraalsest ja lipiidide, valkude ja teiste biomolekulide liigse difusiooni vältimine..
Nagu me defineerisime, rühmitatakse epiteeli rakud rõngasse. See struktuur eraldab raku apikaalse pinna külg- ja basaalsetest, mis määrab domeenide eristamise..
Seda eraldamist peetakse üheks olulisemaks mõisteks epiteeli füsioloogia uurimisel.
Teiseks takistavad tihedad ristmikud ainete vaba liikumist läbi epiteelirakkude kihi, mis muundub paratsellulaarse tee barjääriks..
-Lõika- või vaheühendused

Lõhede või lõhede ühendused on leitud piirkondades, kus naaberrakkude vahel puudub tsütoplasmaatiline membraan. Jagatud ristmikul ühendatakse rakkude tsütoplasmid ja luuakse füüsiline ühendus, kus võib esineda väikeste molekulide läbipääsu..
See liigeste klass on peaaegu kõigis epiteelides ja muudes kudedes, kus need on üsna erinevad..
Näiteks võivad mitmesugustes kudedes rakuvälised signaalid avada või sulgeda, nagu see on neurotransmitteri dopamiini puhul. Selle molekuli olemasolu vähendab võrgusilma klassi neuronite vahelist suhtlust vastuseks valguse suurenenud intensiivsusele.
Lõhustumise liigestes osalevad valgud
Piluühendusi moodustavad valgud, mida nimetatakse konnexiinideks. Seega saadakse "conexón" kuue connexini monomeeni liiduga. See struktuur on õõnes silinder, mis läbib tsütoplasma membraani.
Ühendused on paigutatud nii, et külgnevate rakkude tsütoplasmide vahel luuakse kanal. Lisaks kipuvad connexonid agregeeruma ja moodustama teatud tüüpi plaate.
Piluühenduste funktsioonid
Tänu nende sidemete moodustumisele võib esineda teatud molekulide liikumine naaberrakkude vahel. Transporditava molekuli suurus on määrav, optimaalne läbimõõt on 1,2, näiteks kaltsiumioonid ja tsükliline adenosiinmonofosfaat.
Konkreetselt võib anorgaanilised ioonid ja vees lahustuvad molekulid raku tsütoplasmast üle viia külgnevasse tsütoplasmasse.
Kaltsiumi kontsentratsioonil on selles kanalis oluline roll. Kui kaltsiumi kontsentratsioon suureneb, kalduvad aksiaalsed kanalid sulguma.
Sel viisil osalevad piluühendused aktiivselt rakkude elektrilise ja keemilise ühendamise protsessis, nagu toimub südamelihasrakkudes, mis vastutavad elektriliste impulsside edastamise eest..
-Ankurdamine või ühendused

Kitsaste liigeste all leiame ankurdusliited. Üldiselt asuvad need epiteeli apikaalse pinna läheduses. Selles grupis saame eristada kolme peamist rühma, vööndi zonula adhereni või desmosoomi, makula adhereene või desmosoomipunkti ja desmosoomi..
Sellist tüüpi ristmike korral eraldatakse tsoonide ja adhereenmakulitega ühendatud külgnevad rakumembraanid suhteliselt laiaulatusliku kaugusega - kui me võrdleme neid minimaalse ruumi, mis on kitsaste ristmike puhul..
Intercellulaarne ruum on hõivatud valkudega, mis kuuluvad kadheriinide, desmogleiinide ja desmocoliinide perekonda, mis on seotud tsütoplasmaatiliste plaatidega, millel on teised desmoplaquina, placoglobina ja placofilina valgud..
Kinnituspunktide liigitus
Zonula adherens
Nagu kitsaste liigendite puhul, jälgime ankurdusliidetes ka paigutuse mustrit rõnga või vöö kujul. Zonula adherens on seotud aktini mikrofilmiga kahe valgu - kadheriinide ja kateniinide - koostoime kaudu..
Macula adherens
Mõningatel juhtudel on see struktuur tuntud lihtsalt desmosoomina, see on läbistav ühendus, mis on seotud keratiinist moodustatud vahefilamentidega. Selles kontekstis nimetatakse nimetatud keratiini struktuure "tonofilimanetos". Filamentkiud ulatuvad ühest punktist teise epiteelirakkudes.
Punkt desmosoomid
Need tagavad tugevuse ja jäikuse epiteelirakkudele. Seega arvatakse, et selle peamine funktsioon on seotud külgnevate rakkude tugevdamisega ja stabiliseerimisega.
Desmosoome saab võrrelda teatud tüüpi neetide või keevisõmblustega, kuna need sarnanevad eraldi väikestele kohtadele ja mitte pidevatele ribadele.
Me leiame seda tüüpi liigesed, mis paiknevad vaheldumisi kettaid, mis seovad kardiomüotsüütidesse südamelihas ja ajukestas vooder välispinna aju ja seljaaju.
-Hemidesmosoomid

Hemidesmosoomid kuuluvad asümmeetriliste ristmike kategooriasse. Nende struktuuride ülesandeks on kinnitada epiteeliraku basaaldomeen aluseks oleva baaslaminaadiga.
Terminit hemidesmosome kasutatakse, sest see struktuur tundub sõna otseses mõttes "keskmise" desmosoomina. Kuid nende biokeemilise koostise seisukohast on mõlemad ametiühingud täiesti erinevad.
Tähtis on selgitada, et desmosoomid vastutavad naaberkärje kleepumise eest teise, samas kui hemidesmosoomi funktsiooniks on rakuga liitumine basaallamiga..
Erinevalt adherens macula või desmosome, hemidesmosoomide on erinev struktuur, kuhu kuuluvad: tsütoplasma lehest seostatakse vaheühendi filamendid ja plaat välismembraane, mis vastutab kinnitamiseks hemidesmosome koos Basaalkiht kaudu ankurdamine hõõglambi.
Üks funktsioon hemidesmosoomidest suurendada üldist stabiilsust epiteelikudedele tänu juuresolekul tsütoskeleti vaheühendi filamentide kinnitatud komponentide basaalmembraani.
Rakkude ristmikud taimedes
Taimariigist puudub enamik eespool kirjeldatud raku ristmikke, välja arvatud funktsionaalne vaste, mis sarnaneb piluühendustega.
Taimedes on külgnevate rakkude tsütoplasmid ühendatud teedega või kanalitega, mida nimetatakse plasmodesmseks.
See struktuur loob järjepidevuse ühest taimerakust teise. Kuigi see erineb struktuurselt piluühendustest, on neil väga sarnased rollid, mis võimaldavad väikeste ioonide ja molekulide läbimist.
Meditsiiniline perspektiiv
Meditsiini seisukohast on rakuühistud oluline küsimus. On leitud, et ristumistes osalevaid valke kodeerivate geenide mutatsioonid teisendatakse kliinilisteks patoloogiateks.
Näiteks kui on olemas teatud mutatsiooni kodeeriv geen konkreetset tüüpi claudin (üks neist valkudest, mis vahendavad vastastikmõju -ühingud kitsa tüüp) põhjustab haruldane haigus inimese.
See on neeru magneesiumi kadu sündroom ja sümptomiteks on väike kogus magneesiumi ja krampe.
Lisaks on leitud, et valgu nektiini 1 kodeeriva geeni mutatsioon on vastutav suulaeluse sündroomi või lõhestava huule eest. Seda seisundit peetakse vastsündinute üheks kõige levinumaks väärarenguks.
Mutatsioonid nectin 1 on seostatud ka teiste seisund, mida nimetatakse ectodermal düsplaasia mõjutavad naha, juuste, küünte ja hammastega inimese.
Pemphigus foliaceus on villiline naha patoloogia määrati autoantikehad desmoglein 1, võtmeelemendiks mis on üle pidav kohesiivsus epidermis.
Viited
- Alberts, B., Bray, D., Hopkin, K., Johnson, A.D., Lewis, J., Raff, M., ... & Walter, P. (2015). Oluline rakubioloogia. Garland Science.
- Cooper, G. M., & Hausman, R.E.. Rakk: molekulaarne lähenemine. Sinauer Associates.
- Curtis, H., & Barnes, N. S. (1994). Kutse bioloogiasse. Macmillan.
- Hill, R.W., Wyse, G. A., Anderson, M., & Anderson, M. (2004). Loomade füsioloogia. Sinauer Associates.
- Karp, G. (2009). Rakkude ja molekulaarbioloogia: mõisted ja katsed. John Wiley & Sons.
- Kierszenbaum, A., & Tres, L. (2016). Histoloogia ja rakubioloogia: patoloogia tutvustus. Elsevier Brasiilia.
- Lodish, H., Berk, A., Darnell, J.E., Kaiser, C. A., Krieger, M., Scott, M.P., ... & Matsudaira, P. (2008). Molekulaarrakkude bioloogia. Macmillan.
- Voet, D., & Voet, J. G. (2006). Biokeemia. Ed. Panamericana Medical.