Jaga:



Mis on monofüütiline rühm?
A monofüütiline rühm on liikide rühm, mis on omavahel seotud ainulaadse päritolu, st esivanemate ja kõigi järglaste ajaloo kaudu..
See termin tähistab siis loomulikku rühma. Ta on vastu terminitele polüfüütiline ja parafüütiline. Viimased määratlevad kunstlikud rühmad ebatäieliku (parafüütilise) või erinevate esivanemate järeltulijate (polüfüütiliste) kaasamiseks..
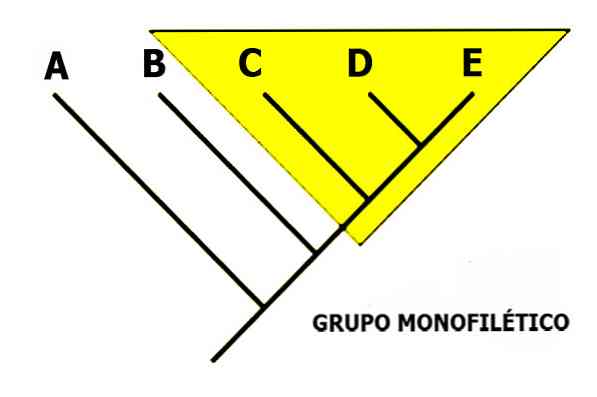
Mõned autorid väidavad, et ainsad loomulikud rühmitused peaksid olema ainult mofiilsed rühmad. Seda seisukohta ei jaga siiski kõik taksonoomid ja süstemaatikud. Numbriline taksonoomia näiteks ei erista mono, para või polüfüütilist taksonit.
Indeks
- 1 Organismide klassifikatsioon
- 2 Taksonoomilised koolid
- 2.1. Numbriline või fenetiline taksonoomia
- 2.2 Evolutsiooniline taksonoomia
- 2.3 Fenogeneetiline või kladistlik taksonoomia
- 3 Koolide vastuolud
- 3.1 Erinevused
- 4 Mõned põhimõisted
- 5 Graafilised kujutised taksonoomiliste koolide järgi
- 5.1 Cladograma
- 5.2 Fenogramm
- 5.3 Fylogramm või füütiline puu
- 6 Viited
Organismide klassifitseerimine
Taksonoomia on teadus, mis vastutab elusolendite klassifitseerimise eest. Selle kohaselt tuleb organismid rühmitada taksoniteks, mis on teineteist välistavad.
Need taksonid jagunevad omakorda kõrgema taseme taksoniteks, mis on üksteist välistavad iga sellise taseme või taksonoomilise kategooria jaoks.
Igas taksonis on organismidel atribuudid (tähemärgid), millele taksonoomid põhinevad, et näidata nende suhteid teiste organismidega ja piiritleda seega bioloogilised taksonid.
Nende märkide vahel esinevate sarnasuste (või erinevuste) hindamiseks ja kaalumiseks on olemas erinevad lähenemisviisid (või koolid) ja tehakse vastavad otsused.
Taksonoomilised koolid
Praegu on kolm peamist taksonoomilist kooli:
Numbriline või fenetiline taksonoomia
R.R. Sokal ja P.H.A. Sneath 1963. aastal. See põhineb vaadeldavate tähemärkide sarnasusel või erinevusel, ilma et arvestataks varasemate hüpoteesidega nende filogeenide kohta organismide klassifitseerimiseks..
Kõigil tähemärkidel on sama väärtus (globaalne sarnasus), võtmata arvesse, kas sarnasused tulenevad homoloogidest või homoplasiatest.
Evolutsiooniline taksonoomia
Seda nimetatakse ka traditsiooniliseks või Darwini taksonoomiaks. Ta kasutab filogeneetilisi suhteid, vanemate ja järglaste suhteid (järjestikused järglased) ning evolutsioonilise muutuse määra organismide liigitamiseks..
Võimaldab gruppidel ema-taksonitest välja jätta, arvestades parafüütilisi taksoneid.
Filogeneetiline või kladistlik taksonoomia
Willi Hennigi poolt 1966. aastal välja pakutud raamatus Filogeneetiline süstemaatika. See põhineb ühistel tuletatud sarnasustel (homoloogiatel) või sünapomorfiatel, et luua evolutsioonilised suhted organismide vahel.
See on aluseks kõige kaasaegsematele bioloogilise klassifikatsiooni süsteemidele ja püüab rühmitada organisme oma evolutsiooniliste suhete kaudu. Ainult tunnustab monofüütiliste rühmade kehtivust.
Koolide vahelised vastuolud
Fenetilise taksonoomia järgib praegu oma kitsas tähenduses väga vähesed taksonoomid, kuid selle tööriistu kasutab sageli ükskõik kumb kahest taksonoomikakoolist..
Damien Auberti sõnul on süstemaatilise taksonoomia praktikat liiga palju aastaid takistanud selle distsipliini aluste põhjalikud erinevused..
Erinevused
Elusolendite adekvaatse klassifitseerimise puhul tuleks esitada teavet, mis tuleks lisada või välja jätta. Kuigi kaks peamist süstemaatikakooli tunnustavad evolutsiooni, on neil vastupidised ideed.
Kladism näitab, et klassifikatsioon peaks peegeldama ainult järjekorda, milles lineaarsete tagajärgede ilmnemine elupuus.
Evolutsioonism omalt poolt väidab, et tuleb arvesse võtta ka filiaalide pikkusena kajastatud muutuste astet. Kooli sõnul kajastaks see pikkus makromajanduslikke katkestusi.
Kladistlik kool väidab, et ükski nende esivanemaid sisaldava rühma järeltulija ei tohiks välja jätta. Teisest küljest nõuab evolutsiooniline taksonoomia selgesõnaliselt, et nende esivanematest väga erinevad järeltulijad tuleks lisada eraldi rühmadesse.
Nii kasutavad mõlemad koolid sageli erinevaid ideid, näiteks „monofiilseid“. See asjaolu, Aubert'i sõnul, muudab filogeneetika uuringud ülemaailmselt ebaühtlaseks ja seetõttu on taksonoomiline liigitus väga ebastabiilne.
Lõpuks saame järeldada, et kui me tahame teha analüüsi ühe või mitme taksoni liigitamiseks ja kasutada kolme kooli postulaate eraldi, on tulemused tõenäoliselt erinevad.
Mõned põhimõisted
Monofüütilise mõiste nõuetekohaseks mõistmiseks peame kladistliku kooli järgi kasutama teatud põhiterminoloogiat:
Märk: mis tahes täheldatav omadus organismis, mille erinevaid ilminguid nimetatakse näiteks juuksed, suled või kaalud; geograafiline jaotus; käitumine jne.
Isiku seisund: iga viis, kuidas seda iseloomu võib esitada, kas primitiivne või tuletatud. Näiteks on inimeste kahepoolne jalutamine tuletatud seisund (iseloom), mitte aga teiste hominiidide neljas otsas (seisund või esivanem)..
Plesiomorfne iseloom: kogu monofüütilise grupi jagatud primitiivne või esivanem iseloom.
Simplesiomórfia: plesiomorphy jagatud kahe või enama taksoniga.
Tuletatud või apomorfne iseloom: kas see tuleneb esivanemast, see tähendab, et see tuleneb uuritava grupi iseloomu muutumisest. See on uue kladi algus.
Autapomorfia: tuletatud mittejagatud iseloom. See esineb ainult taksonis ja seda kasutatakse sageli mikrotaksonoomias, et eristada liike.
Sinapomorfia: kahe või enama liigi või taksoni jagatud apomorfia või iseloom.
Clado (monofiletico): rühm, mis koosneb esivanemate liikidest ja kõigist selle järeltulijatest.
Homoloogia: sarnasuse tingimus ühise esivanema olemasolu tõttu.
Homoloogne iseloom: sarnased tähemärgid või erinevad atribuudid, kuid mis pärinevad ühisest esivanemast.
Analoogia: sarnaste struktuuride arendamine, mis täidavad sama funktsiooni, kuid nende embrüonaalne päritolu on erinev.
Homoplasy: vale sarnasus, mis tuleneb erinevate esivanemate tähemärkide olemasolust. Esineb lähenemise, paralleelsuse või ümberpööramisega.
Lähenemine: on analoogia sünonüüm.
Paralleelsus: sama iseloomu sõltumatu areng samast esivanema iseloomust.
Pöördumine: apomorfia, mis kaotatakse hiljem (naaseb plesiomorfsesse seisundisse) ühes monofüütilise rühma taksonites.
Graafilised esitusviisid taksonoomiliste koolide järgi
Cladograma
Kladogramm on kladistliku kooli iseloomulik diagramm. Nendes väljendatakse genealoogilisi filogeneetilisi suhteid, mis peavad olema loomulikud või monofülilised, see tähendab, et nad sisaldavad ühist esivanemat ja tema järeltulijaid.
Fenogramm
Fenogrammid on skeemid, mida füsioloogiline taksonoomia kasutab organismide liigitamiseks. Selline analüüs võtab vastu kolme taksonitüüpi: monofüütiline, parafüütiline ja polüfüütiline.
Kuigi need diagrammid on suhteliselt sarnased kladogrammidega, ei väljenda need filogeneetilisi suhteid, vaid pigem sarnasust või nähtavat erinevust organismide vahel..
Fylogramm või füütiline puu
Evolutsioonilise või klassikalise taksonoomilise kooli pakutud filogeneetilised klassifikatsioonid kasutavad füütilisi puid. Need diagrammid väljendavad järglaste esivanemate genealoogilisi suhteid ja aktsepteerivad kahte tüüpi taksoneid: monofüütiline ja parafüütiline.

Viited
- D. Aubert (2015). Filogeneetilise terminoloogia ametlik analüüs: Süstemaatika praeguse paradigma läbivaatamine. Phytoneuron
- D. Baum (2008). Filogeneetilise puu lugemine: monofüütiliste rühmade tähendus. Loodusharidus
- L.M. Chiappe & G. Dyke (2002). Lindude mesosooosne kiirgus. Ökoloogia ja süstemaatika iga-aastane ülevaade.
- Cladistics Wikipedias. Välja otsitud andmebaasist: en.wikipedia.org/wiki/Cladistics
- W. Hennig (1966). Phylogenetic Systematic. Illinois Press, Urbana
- Monofeed Wikipedias. Välja otsitud andmebaasist: en.wikipedia.org/wiki/Monophyly
- P.A. Reeves & C.M. Richards (2007). Terminali monofüütiliste rühmade eristamine retikuliseeritud taksonitest: Fenetiliste, puidupõhiste ja võrgu protseduuride toimimine. Süstemaatiline bioloogia