Jaga:



Pentoosifaaside ja nendega seotud haiguste teed
The pentooside tee Fosfaat, tuntud ka kui heksooside monofosfaadi kõrvalekalle, on põhiline metaboolne rada, millel on lõpp-produktina ribosoomid, mis on vajalikud nukleotiid- ja nukleiinhappe sünteesiteedel, nagu DNA, RNA, ATP, NADH, FAD ja koensüüm A.
Samuti toodab see NADPH-d (nikotiinamiidadeniindinukleotiidfosfaati), mida kasutatakse mitmesugustes ensümaatilistes reaktsioonides. See marsruut on väga dünaamiline ja suudab oma tooteid kohandada sõltuvalt rakkude hetkevajadustest.
ATP (adenosiintrifosfaat) loetakse raku "energiavaluutaks", sest selle hüdrolüüsi saab siduda paljude biokeemiliste reaktsioonidega.
Samamoodi on NADPH teine oluline energiavaluuta rasvhapete redutseerivaks sünteesiks, kolesterooli sünteesiks, neurotransmitterite sünteesiks, fotosünteesiks ja detoksikatsioonireaktsioonideks..
Kuigi NADPH ja NADH on struktuuris sarnased, ei saa neid biokeemilistes reaktsioonides kasutada vaheldumisi. NADPH osaleb vaba energia kasutamisel teatud metaboliitide oksüdeerimisel redutseerivaks biosünteesiks.
Seevastu on NADH seotud vaba energia kasutamisega metaboliitide oksüdatsioonist ATP sünteesimiseks.
Indeks
- 1 Ajalugu ja asukoht
- 2 Funktsioonid
- 3 faasi
- 3.1 Oksüdeeriv faas
- 3.2 Mitteoksüdatiivne faas
- 4 Seotud haigused
- 5 Viited
Ajalugu ja asukoht
Selle marsruudi olemasolu märked algasid 1930. aastal tänu uurijale Otto Warburgile, kellele NADP avastamine talle omistati+.
Teatud tähelepanekud võimaldasid marsruudi avastamist, eriti hingamise jätkamist glükolüüsi inhibiitorite, näiteks fluoriidi ioonide juuresolekul..
Seejärel kirjeldasid pentoosfosfaadi rada 1950. aastal teadlased Frank Dickens, Bernard Horecker, Fritz Lipmann ja Efraim Racker.
Kolesterooli ja rasvhapete sünteesiga seotud kudedel, näiteks piimanäärmetel, rasvkoes ja neerudes, on kõrge pentoosfosfaadi ensüümide kontsentratsioon.
Maksa on samuti selle tee jaoks oluline koe: umbes 30% glükoosi oksüdatsioonist selles koes toimub tänu pentoosfosfaadi raja ensüümidele..
Funktsioonid
Pentoosfosfaadi rada vastutab süsiniku homöostaasi säilitamise eest rakus. Samamoodi sünteesib rada aminohapete sünteesiga seotud nukleotiidide ja molekulide (peptiidide ja valkude struktuursete plokkide) prekursorid.
See on ensüümreaktsioonide vähendamise peamine allikas. Lisaks annab see vajalikud molekulid anaboolsete reaktsioonide ja oksüdatiivse stressi vastaste kaitseprotsesside jaoks. Raja viimane etapp on kriitilise tähtsusega kriitilistes olukordades.
Faasid
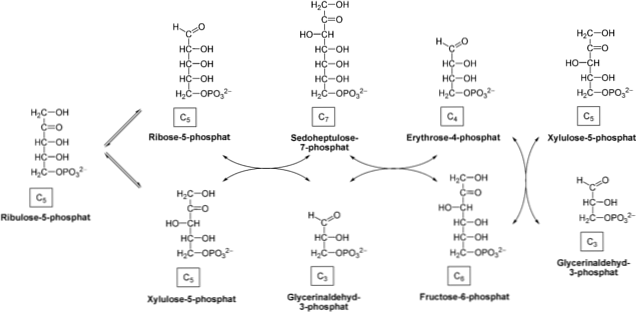
Pentoosfosfaadi rada koosneb rakulise tsütosooli kahest faasist: oksüdatiivsest, mis genereerib NADPH glükoosi-6-fosfaadi oksüdeerimise teel riboos-5-fosfaadiks; ja mitteoksüdatiivne, mis hõlmab kolme, nelja, viie, kuue ja seitsme süsiniku suhkrute muundamist.
Sellel marsruutil on ühised reaktsioonid Calvini tsükli ja Entner-Doudoroffi rajaga, mis on alternatiiv glükolüüsile.
Oksüdeeriv faas
Oksüdatiivne faas algab glükoosi-6-fosfaatmolekuli dehüdrogeenimisega süsinikul 1. Seda reaktsiooni katalüüsib glükoosi-6-fosfaadi dehüdrogenaasi ensüüm, millel on NADP-ga kõrge spetsiifilisus.+.
Selle reaktsiooni saadus on 6-fosfonoglükono-5-laktoon. Seejärel hüdrolüüsib see produkt ensüüm laktonaas, et saada 6-fosfoglükonaat. Viimane ühend on võetud 6-fosfoglükonaadi dehüdrogenaasi ensüümiga ja muutub ribuloos-5-fosfaadiks.
Fosforpentoos-isomeraas ensüüm katalüüsib oksüdatiivse faasi viimast etappi, mis hõlmab riboosi 5-fosfaadi sünteesi ribuloos-5-fosfaadi isomerisatsiooni teel..
See reaktsioonide seeria tekitab kaks NADPH molekuli ja ühe riboos-5-fosfaadi molekuli glükoosi 6-fosfaadi molekuli kohta, mis siseneb sellesse ensümaatilisse rada..
Mõnedes rakkudes on NADPH nõuded suuremad kui riboos-5-fosfaatidel. Seetõttu võtavad transketolaasi ja transaldolaasi ensüümid riboso-5-fosfaati ja muudavad selle glütseraldehüüdi 3-fosfaadiks ja fruktoos-6-fosfaadiks, andes tee mitte-oksüdatiivsele faasile. Need kaks viimast ühendit võivad siseneda glükolüütilise rada.

Mitteoksüdatiivne faas
Faas algab epimerisatsioonireaktsiooniga, mida katalüüsib ensüüm pentoos-5-fosfaadi epimeraas. See ensüüm võtab ribuloos-5-fosfaadi ja muundub ksüluloos-5-fosfaadiks.
Toode võtab üle transketolaasi ensüüm, mis toimib koos koensüümi tiamiinpürofosfaadiga (TTP), mis katalüüsib ksüluloos-5-fosfaadi läbimist riboos-5-fosfaadile. Ketoosi ülekandmisel aldoosile valmistatakse glütseraldehüüd-3-fosfaat ja sedoheptuloos-7-fosfaat..
Järgnevalt kannab transaldolaasi ensüüm C3 sedoheptuloos-7-fosfaadi molekulilt glütseraldehüüd-3-fosfaadile, mis toodab nelja süsiniku suhkrut (erütroos-4-fosfaati) ja kuue süsiniku suhkrut (fruktoosi-6). -fosfaat). Need tooted on võimelised toitma glükolüütilist rada.
Transketosala ensüüm toimib uuesti, et viia ksüluloos-5-fosfaadi C2 erütroos-4-fosfaadile, mille tulemuseks on fruktoos-6-fosfaat ja glütseraldehüüd-3-fosfaat. Nagu eelmises etapis, võivad need tooted siseneda glükolüüsi.
See teine etapp ühendab NADPH-d genereerivad teed nendega, kes vastutavad ATP ja NADH sünteesi eest. Lisaks võivad fruktoos-6-fosfaadi ja glütseraldehüüdi-3-fosfaadi produktid siseneda glükoneogeneesi..
Seotud haigused
Erinevad patoloogiad on seotud pentoosfosfaadi rajaga, nende neuromuskulaarsete haiguste ja erinevate vähivormide vahel.
Enamik kliinilisi uuringuid keskenduvad glükoosi-6-fosfaadi dehüdrogenaasi aktiivsuse kvantifitseerimisele, sest see on peamine ensüüm, mis vastutab raja reguleerimise eest.
Aneemia suhtes vastuvõtlikele vererakkudele on neil glükoosi-6-fosfaadi dehüdrogenaasi ensümaatiline aktiivsus madal. Seevastu kõri kantseroomidega seotud rakuliinidel on kõrge ensüümi aktiivsus.
NADPH osaleb glutatiooni tootmisel, mis on võtmetähtsusega peptiidmolekul, mis kaitseb oksüdatiivset stressi..
Erinevad vähivormid viivad pentoosirada aktiveerumiseni ja on seotud metastaaside, angiogeneesi ja kemoteraapia ja kiiritusravi ravivastustega..
Teisest küljest areneb krooniline granulomatoosne haigus, kui NADPH tootmisel esineb puudusi.
Viited
- Berg, J.M., Tymoczko, J.L., Stryer, L (2002). Biokeemia. WH Freeman
- Konagaya, M., Konagaya, Y., Horikawa, H., & Iida, M. (1990). Pentoosfosfaadi rada neuromuskulaarsetes haigustes - lihasglükoosi 6-fosfaadi dehüdrogenaasi aktiivsuse ja RNA sisalduse hindamine. Rinsho shinkeigak. Kliiniline neuroloogia, 30(10), 1078-1083.
- Kowalik, M.A., Columbano, A., & Perra, A. (2017). Pentoosfosfaadi raja kujunemine hepatotsellulaarses kartsinoomas. Onkoloogia piirid, 7, 87.
- Patra, K. C. ja Hay, N. (2014). Pentoosfosfaadi rada ja vähk. Biokeemiliste teaduste suundumused, 39(8), 347-354.
- Stincone, A., Prigione, A., Cramer, T., Wamelink, M., Campbell, K., Cheung, E., ... & Keller, M. A. (2015). Metabolismi taastumine: pentoosfosfaadi raja biokeemia ja füsioloogia. Bioloogilised ülevaated, 90(3), 927-963.
- Voet, D., & Voet, J. G. (2013). Biokeemia. Artmed Publisher.