Jaga:



Foraminifera omadused, morfoloogia, klassifikatsioon, elutsükkel
The foraminifera nad on amoeboidi algloomade, mõnede mere- ja teiste mageveekogude grupp. Nad ilmusid primaarse ajastu alguses (Kambrium) ja nende järeltulijad on asustanud praegused ookeanid. Neid võib leida rannikualadest (hüpo- või hüpersaliin) ookeani põrandale ning troopikatest külma Arktika ja Antarktika ookeanide poole.
Selle jaotus sõltub mitmest tegurist. Näiteks on olemas liike, mis toetavad suuri ja sagedasi temperatuurimuutusi, samas kui teised ei suuda seda ellu jääda, nii et ookeanide termiline struktuur tähistab olulisi erinevusi foraminifera ühenduste vahel..

Samuti on foraminifera jaotuse määravaks teguriks ka sügavus nende otsese esinemise tõttu valguse tungimisel. Teisest küljest on rõhk seotud eelnevate teguritega (temperatuur ja sügavus), mis mõjutavad otseselt CO lahustuvust2, mis mõjutab kaltsiumkarbonaadi sekretsiooni kestade moodustamiseks.
Teisest küljest on madalates piirkondades vee energia oluline element, kuna see mõjutab substraadi tüüpi (kõva või pehme) ja toitainete jaotumist..
Samamoodi võivad teised faktorid, nagu soolsus, vee hägusus, pH, mikroelementide olemasolu ja / või orgaanilised komponendid, voolud, settimise määr, kindlaks määrata kohapeal foraminifera jaotumise..
Indeks
- 1 Tuntud liigid
- 2 Omadused
- 2.1 Foraminifera suuruse ja morfoloogiaga seotud tegurid
- 3 Taksonoomia
- 4 Klassifikatsioon
- 4.1 Athalamea
- 4.2 Monotalamea
- 4.3 Ksenofoofia
- 4.4 Tubotalamea
- 4.5 Globotalamea
- 5 Morfoloogia
- 5.1 -Suurus
- 5.2 -Protoplasma
- 5.3 - luustik või kest
- 5.4 - Foraminifera liigid
- 5.5 -Seudopoodid
- 6 Elutsükkel
- 7 Paljundamine
- 8 Toitumine
- 9 Rakendused
- 9.1 Mauritiuse juhtum
- 10 Viited
Tuntud liigid
Praegu on teada rohkem kui 10 000 liiki, samas kui umbes 40 000 on kadunud. Mõnedel liikidel on merepõhja elupaigad, see tähendab, et need on põhjaorganismid, kes elavad sageli epifauna (epibetónicos) osaks oleval liival, või elavad liiva all (endobetónicos). Seetõttu tuntakse neid ka elava liivana.
Nad võivad elada ka taimedel, kus nad puhuvad epifüütidena ja isegi paljud neist valivad vaikset elu, see tähendab, et nad elavad aluspinna külge kogu nende olemasolu vältel..
Samal ajal kui teised foraminifera elavad ookeanis erinevatel sügavustel (0 kuni 300 m), see tähendab, et nad on planktoniline elu, mis moodustab osa mere mikroplanktonist. Need vormid on napid ja vähem mitmekesised.
Suurim ja kõige keerulisem planktoniline foraminifera on tavalisem troopilistes ja subtroopilistes keskkondades. Kõrgetel laiuskraadidel on need organismid tavaliselt väikesed, väiksemad ja väga lihtsal viisil.
Omadused
Foraminifera iseloomustav omadus on karkass või kest, struktuur, mis on võimaldanud välja surnud vorme uurida mere mikroflooride kujul, mis ladestatakse mere põhja..
Seega on foraminifera diferentseerimiseks põhiline element ja see on organismi ainuke struktuur, mis fossiilib. Need fossiilid on mere settes väga rikkalikud, osalevad ka settekivimite moodustamisel.
Kooride peamised keemilised ühendid on kaltsiit, aragoniit ja ränidioksiid. Embrüokambri kuju ja mõõtmed sõltuvad selle päritolust, olenemata sellest, kas tegemist on seksuaalse või ebatavalise reprodutseerimisega.
Onogeensuse ajal kontrollib foraminifera kambrite kasvu ja suurust. See kontroll toimub pseudopoodiliste voolude pikkuse ja paigutuse kaudu, kuna pseudopoodid vastutavad mineraalsele kestale eelneva orgaanilise membraani moodustumise eest..
See protsess on rakuprotsesside säilitamiseks väga oluline, kuna kaamera toimib bioreaktorina.
Tegurid, mis mõjutavad foraminifera suurust ja morfoloogiat
Tuleb märkida, et foraminifera kasutatav suurus ja lõplik morfoloogia sõltuvad mitmest tegurist, sealhulgas:
- Embrüokambri kuju ja mõõtmed.
- Kasvamisetappide arv täiskasvanu staadiumisse (st foraminifera kambrite arv).
- Kaamera kuju ja selle muudatused ontogeeni ajal.
- Kaamerate paigutus.
Suurematel foraminifera-del on strateegilised kasvumudelid, et säilitada konstantsete kambrite maht optimaalset suurust ületamata. Need strateegiad seisnevad kaamerate jagamises mitmesse kambrisse, mida nimetatakse klikkideks.
Need klastrid on paigutatud nii, et oleks tagatud transpordi- ja regulatsioonifunktsioonid kambrite sisemuses oleva protoplaasi ja väljastpoolt. See tähendab, et kõik kaamerad ja klipid on omavahel täielikult ühendatud.
Kambrite paigutus võib toimuda sirgjoonelise või spiraalse teljega. See sõltub pseudopoodiliste voolude asukohast ja ava või avade asukohast kambris.
Taksonoomia
Valdkond: Eukarya
Kuningriik: Protista
Ilma Range: SAR Supergroup
Superphylum: Rhizaria
Varjupaik: Foraminifera
Klassid ja tellimused
- Athalamea (Reticulomyxida)
- Monothalamea (Allogromiida, Astrorhizida, Komokiida)
- Ksenofoofia (psamminiid, stannomiid)
- Tubotalamea(Fusulinida, Involutinida, Milioliid, Silicoloculinide, Spirilliniid)
- Globothalamea(Lituolida, Loftusiida, Schlumbergerinida, Textulariida, Trochaminminida, Rotalide, Buliminide, Globigerinide, Robertinida, Carterinide, Lagenida).
Klassifikatsioon
Kuigi on veel palju selgitusi, on seni võimalik eristada 5 klassi:
Athalamea
Siit leiate foraminifera, millel ei ole kesta või mis on alasti.
Monotalamea
See hõlmab betoonset foraminifera, mis sisaldab ühe kambriga orgaanilist või aglutineeritud koort.
Ksenofoofia
Sel juhul on foraminifera spetsialiseerunud betoonitüübiga, suure suurusega, mitmekihiline ja aglutineeritud koorega. Tavaliselt on need detritivoorid või saprofaadid, st nad saavad oma toidu detriidist või orgaanilisest ainest lagunemisel..
Tubotalamea
See hõlmab betoonilist foraminifera, millel on vähemalt alaealiste etapil mitu torukambrit, mida saab spiraalina haavata, sidestatud või lubjatu koorega..
Globothalamea
See klassifikatsioon hõlmab nii betoonseid kui ka planktonilisi foraminiferaid, mis sisaldavad globaalset, aglutineeritud või lubjakest vormi. Koored võivad olla uniseriate, biseriada, triseriada või trocospiraladas.
See klassifikatsioon on aga pidevas arengus.
Morfoloogia
-Suurus
Foraminifera suurus on tavaliselt vahemikus 0,1 kuni 0,5 cm, mõnel liigil on võimalik mõõta 100 μm kuni 20 cm..
-Protoplasm
Foraminifera moodustavad protoplasmaatiline mass, mis moodustab foraminifera raku.
Protoplasm on tavaliselt värvitu, kuid võib mõnikord sisaldada väikeses koguses orgaanilisi pigmente, lipiide, sümbiootilisi vetikaid või rauaühendeid, mis annavad selle värvuse.
Protoplasm koosneb sisemisest osast, mida nimetatakse endoplasmaks ja väliseks osaks ektoplasmaks.
Endoplasmas on see kaitstud koorega ja selles jagunevad organellid seedetrakti vacuoles, tuumas, mitokondrites, graanulites, Golgi seadmetes või ribosoomides. Seetõttu nimetatakse seda mõnikord granuleeritud endoplasmaks. Ektoplasm on läbipaistev ja sealt lahkuvad sissetõmmatavad pseudopoodid.
Protoplasma piirneb väliselt orgaanilise membraaniga, mis koosneb pealekantavatest mukopolüsahhariidilehtedest..
Protoplasmaatiline mass laieneb koorest läbi ühe või mitme avause (pooride) ja katab selle väliselt (kaameraväline protoplasm) ja see moodustab pseudopoodia..
-Luustik või kest
Foraminifera fikseerib oma rakupinna püsivalt mineraalse karkassi (kest) ehitamise teel..
Kesta moodustavad septa poolt eraldatud kambrid, kuid samal ajal suhtlevad nad üksteisega omavaheliste aukude kaudu, mida nimetatakse foramina, seega ka foraminifera nime. Karkassi või kesta keemiline koostis muudab need struktuurid, mis fossiilivad väga kergesti.
Kambrite sisemus on kaetud kitiiniga väga sarnase orgaanilise materjaliga. Lisaks võib kesta põhilõikudel olla; Te saate ka oma poorid oma väljapoole või puududa.
Mineraalset kihti võib moodustada ühe kambriga (primitiivne foraminifera või monotalamos) või kambris, mis kasvab pidevalt, või mitme kambriga, mis on moodustatud järjestikustes etappides, keerulises katkematu kasvu süsteemis (foraminifera polylamas).
See viimane protsess seisneb eelnevalt moodustatud kesta lisamises ja strateegilistes kohtades uue skeleti materjaliga.
Paljud foraminifera on võimelised valima oma kesta moodustava materjali vastavalt nende keemilisele koostisele, suurusele või kujule, kuna substraadiga kokkupuutuvad marginaalsed pseudopoodilised voolud suudavad seda ära tunda.
-Foraminifera liigid
Kesta konstruktsiooni järgi võib neid klassifitseerida kolme peamisse Foraminifera tüüpi:
Aglutineeritud (või \ t
Sellist tüüpi kestas koguvad foraminifera koos pseudopoodidega suure hulga orgaanilisi materjale, mis on kättesaadavad nende elukeskkonnas, mis seejärel aglutineerub, nagu mineraalsed terad, käsnad, piimad jne..
Enamik aglutineeritud foraminifera tsementeerib oma kesta kaltsiumkarbonaadiga, kuid kui seda ühendit keskkonnas ei esine, näiteks need, mis elavad ookeani sügavates piirkondades, kus kaltsiumi ei eksisteeri, võivad nad seda teha silikoonsete, ferrugiinsete orgaaniliste tsementidega, jne
Portselan
Sellisel juhul moodustatakse kest magneesiumkalsiidi nõeltega, mis sünteesitakse foraminifera Golgi aparaadis..
Need nõelad transporditakse ja kogunevad väljastpoolt ning need võivad olla võõrkonstruktsioonide (tsement) ühenduselemendid või moodustada otse välimine karkass. Neid leidub hüpersaliini keskkonnas (> 35% soolsus).
Nad on tavaliselt imelikud, see tähendab, et neil on tavaliselt pseudo-poorid, mis ei läbi koorikut täielikult.
Hüaliinid
Need moodustuvad kaltsiitkristallide kasvu tõttu tänu orgaanilisele vormile, mis on moodustatud biolineraliseerimisprotsessist (mineralisatsioon in situ), mis viiakse läbi protoplasmaatilisele kehale..
Neile on iseloomulik, et seina paksus on läbipaistev. Samuti on need perforeeritud, kui pooride asukoht, tihedus ja läbimõõt varieerub vastavalt liigile.
-Pseudopoodid
Seda struktuuri kasutatakse mobilisatsiooniks, substraatide kinnitamiseks, saagiks ja skeleti loomiseks. Pseudopoodide tagasitõmbamiseks ja laiendamiseks on foraminifera keerulise mikrotuubulite võrgustik, mis on paigutatud rohkem või vähem paralleelsetesse ridadesse.
Pseudopoodide laiendamine võib ulatuda kaks või kolm korda keha pikkusest ja võib isegi olla kuni 20 korda pikem. See sõltub igast konkreetsest liigist.
Liikumise ajal liikumine on otseselt seotud kesta kuju ja avade asendiga (kus pseudopoodid on pärit).
Kuid enamik foraminifera liigub järgmiselt: pseudopoodid kinnituvad substraadile ja seejärel suruvad ülejäänud rakku. Sel viisil liikumine võib kiirendada umbes 1–2,5 cm / tunnis.
Teisest küljest nimetatakse foraminifera pseudopoode Granurreticulopodiaks, sest pseudopoodide sisemuses on kahesuunaline tsütoplasmaatiline vool, mis kannab graanuleid..
Graanulid võivad koosneda erinevate materjalide osakestest, mitokondritest, seedetraktist või prügi-vaakoolidest, sünbiootilistest delloflagellatidest jne. Sel põhjusel on grupi üks sünonüüme Granuloreticulosa.
Pseudopoodide teine oluline omadus on see, et nad kipuvad olema pikad, õhukesed, hargnenud ja väga rikkad, moodustades sel viisil retikulopoodia võrgustiku virnastamise teel (anastomoos)..
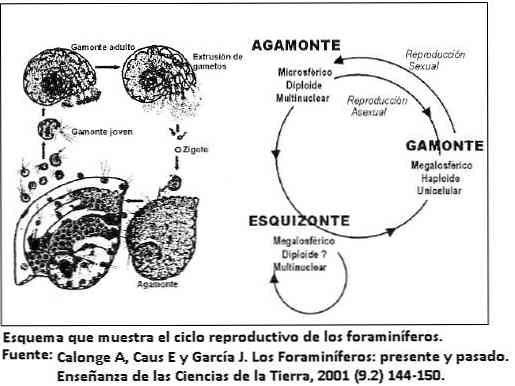
Elutsükkel
Foraminifera elutsükkel on tavaliselt lühike, tavaliselt paar päeva või nädalat, kuid suurtes vormides võib elutsükkel olla kaks aastat.
Kestus sõltub elu strateegiast, mida foraminifera aktsepteerib. Näiteks töötavad väikese kujuga lihtsa morfoloogiaga lühikesed oportunistlikud strateegiad.
Kuna suured vormid ja kooriku erakordselt keeruline morfoloogia arendavad konservatiivset elustrateegiat.
Viimane käitumine on ühekordsete organismide puhul väga haruldane; võimaldab neil säilitada ühtlase rahvastikutiheduse ja aeglase kasvu.
Paljundamine
Enamik foraminifera esineb kahel morfoloogial: põlvkondade vaheldumine vastavalt reproduktsioonitüübile, seksuaalne või ebatavaline, välja arvatud planktoniline foraminifera, mis reprodutseerib ainult seksuaalselt.
Seda morfoloogia muutust nimetatakse dimorfismiks. Tekkinud seksuaalse reproduktsiooni vormi (gamogooniat) nimetatakse gamoniks, samas kui ebatavaline reprodutseerimine (skisogoonia) annab skeemi vormi. Mõlemad on morfoloogiliselt erinevad.
Mõned foraminifera koordineerivad paljunemistsüklit hooajalise tsükliga, et optimeerida ressursside kasutamist. Ei ole ebatavaline, et enne seksuaalse põlvkonna esinemist beetoonsetes vormides esineb mitu pidevat ebatavalist reproduktsiooni.
See seletab, miks skeemivormid on rohkemad kui gamontide vormid. Algselt kujutab gamon ühte tuuma ja jagab seejärel paljude sugurakkude tootmiseks.
Samal ajal kui horisontaaljooned on mitmekihilised ja pärast meioosi fragmente moodustavad uued sugurakud.
Toitumine
Foraminifera iseloomustab heterotroofne, st nad toidavad orgaanilist ainet.
Sel juhul söödavad foraminifera peamiselt diatomeid või baktereid, kuid teised suuremad liigid söövad nematoode ja koorikloomi. Vangid on nende pseudopoodide kaudu lõksus.
Samuti võivad need organismid kasutada mitmesuguste sümbiontide vetikate, nagu rohelised, punased ja kuldvetikad, samuti diatomeid ja dinoflagellate, ning paljudes neist võib olla isegi väga keeruline..
Teisest küljest on mõned foraminifera liigid kleptoplastilised, mis tähendab, et fotosünteesi funktsiooni jätkamiseks muutuvad allaneelatud vetikate kloroplastid osaks foraminiferaadist..
See kujutab endast alternatiivset võimalust elada energiat.
Rakendused
Foraminifera fossiilse rekordi arvukus geoloogilise aja, evolutsiooni, keerukuse ja suuruse tõttu muudab nad lemmikvahendiks Maa praeguse ja mineviku uurimiseks (geoloogiline kella).
Seetõttu on selle suur liikide mitmekesisus väga kasulik biostratigraafilise, paleoökoloogilise, paleokeanograafilise tüübi uuringutes..
Kuid see võib samuti aidata vältida ökoloogilisi katastroofe, mis võivad majandust mõjutada, sest muutused foraminifera populatsioonides viitavad keskkonnamuutustele..
Näiteks foraminifera ilma kooreta, on tundlikud keskkonnamuutustele ja reageerivad kiiresti ümbritseva keskkonna muutustele. Seetõttu on need ideaalsed indikaatorliigid reefivee kvaliteedi ja tervise uurimiseks.
Mauritiuse juhtum
Ka mõned sündmused on meid selle üle mõtisklenud. Niisugune on nähtus Mauritiuses täheldatud nähtuse puhul, kus osa ranniku valgest liivast kadus ja nüüd peavad nad seda Madagaskarilt turismivoo säilitamiseks importima..
Ja mis seal juhtus? Kust pärineb liiv? Miks see kadus??
Vastus on järgmine:
Liiv on midagi enamat kui paljude organismide kaltsiumkarbonaadi kestade kogunemine, nende hulgas ka foraminifera, mis tõmmatakse ranna servale. Liiva kadumine oli tingitud karbonaatide tootjate järkjärgulisest ja püsivast vähenemisest.
See juhtus merede reostamisega lämmastiku ja fosforiga, mis jõuavad rannikutesse, kuna väetised on liigselt kasutatud teatavate toodete, näiteks suhkruroo, külvamisel..
Sellepärast on oluline uurida foraminifera sotsiaalteadustes, et vältida keskkonnakatastroofe, nagu eespool kirjeldatud, mis mõjutavad otseselt majandust ja ühiskonda..
Viited
- Wikipedia toetajad. Foraminifera [online] Vikipeedia, vaba entsüklopeedia, 2018 [konsultatsiooni kuupäev: 1. november 2018]. Saadaval aadressil es.wikipedia.org.
- Calonge A, Caus E ja García J. Foraminifera: kohal ja minevik. Maateaduste õpetamine, 2001 (9,2) 144-150.
- Hromic T. Bioloogiline mitmekesisus ja mikrobentose (Foraminifera: Protozoa) ökoloogia, Boca del Guafo ja Golfo de Penas (43º-46º s), Tšiili vahel. Cienc. Tecnol. 30 (1): 89-103, 2007
- Humphreys AF, Halfar J, Ingle JC jt. Merevee temperatuuri, pH ja toitainete mõju väikese arvukuse madalate veealuste põhjavee foraminifera levikule ja iseloomule Galapagose piirkonnas. PLoS One 2018, 13 (9): e0202746. Avaldatud 2018. sept. 12. doi: 10.1371 / journal.pone.0202746
- De Vargas C, Norris R, Zaninetti L, Gibb SW, Pawlowski J. Molekulaarsed tõendid krüptilisest spekulatsioonist planktonilistes foraminifeerides ja nende seos ookeani provintsidega. Proc Natl Acad Sci. U. A., 1999, 96 (6): 2864-8.